

填写下面的表格,我们将向您发送PDF版本的电子邮件“裂解与溶菌性 - 了解噬菌体生命周期”
噬菌体(噬菌体)是特异性感染细菌的强制性细胞内病毒。他们是由两名研究人员弗雷德里克·威廉·托特(Frederick William Twort)独立发现的11915年在伦敦大学和费利克斯·德·赫雷尔(FélixD’Herelle)2他确认了这一发现,并在1917年创造了噬菌体一词,此后对此进行了大量研究。
噬菌体结构
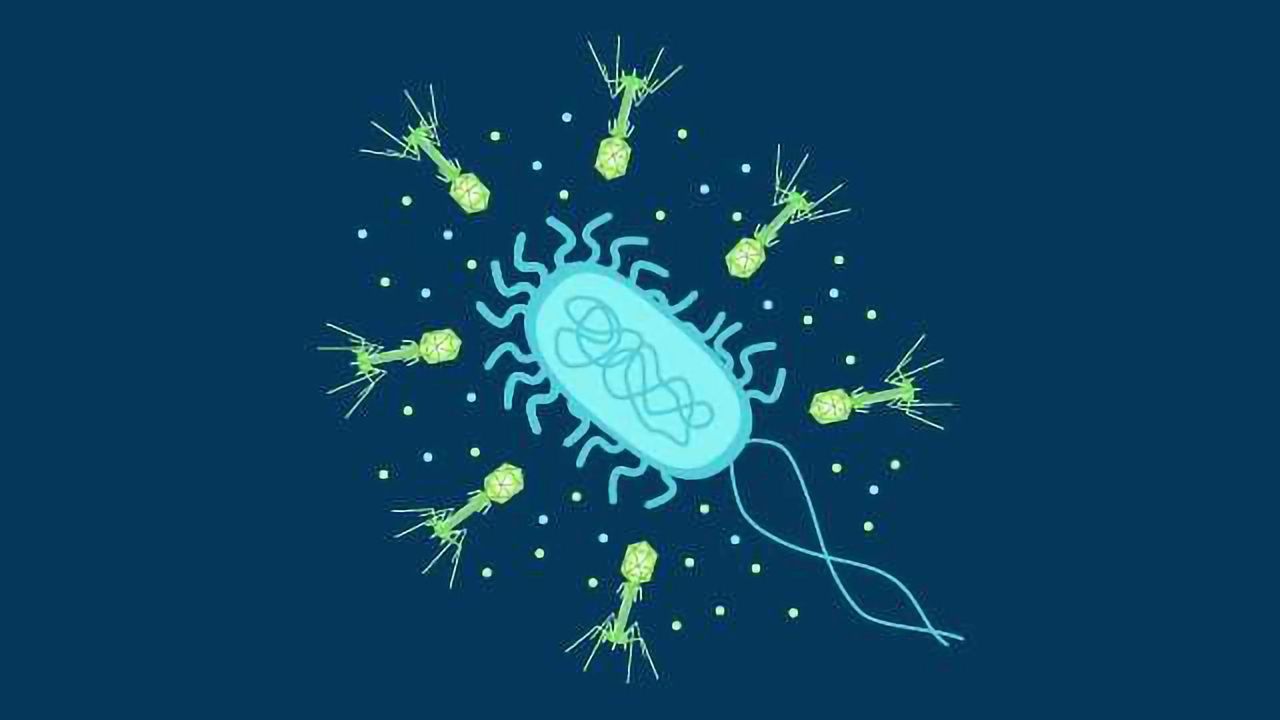
噬菌体具有非常简单的结构(图1)。它们的遗传物质包含在棱镜形状的头部中,周围是蛋白质衣壳。这是通过颈部或项圈区域连接到细长的鞘(有时称为尾巴)的。
鞘形成一个空心管,将病毒DNA/RNA注入宿主细胞中,并被保护性护套蛋白包围。在护套的底部是底板,尾纤维(通常为六个)促进促进宿主电池的附着的底板。

图1。噬菌体的示例结构。
为了复制,噬菌体必须首先进入宿主单元。它们与细菌细胞表面上的特定受体结合其尾纤维(吸附),并创建一个孔,该过程与附着一起由底板协调3。将刚性管从鞘中推出,刺穿细菌细胞膜中的一个孔,通过该孔注入其遗传物质(DNA或RNA,双或单链)。然后,如果周围条件在称为The The The的过程中不利,他们可以劫持主机单元的蜂窝机器以进行自身复制裂解周期。或者,如果条件良好,它们可能会在宿主细胞内进入休眠状态,称为溶酶发生循环。
裂解周期
在裂解周期(图2)中,有时被称为毒性感染,感染噬菌体最终杀死了宿主细胞以产生许多自己的后代。注入宿主细胞后,噬菌体基因组立即合成了分解宿主DNA的早期蛋白质,从而使噬菌体能够控制细胞机械。
然后,噬菌体使用宿主细胞合成构建新的噬菌体颗粒所需的剩余蛋白质。头部和护套分别组装,新的遗传材料堆积在头部,并构建了新的女儿噬菌体颗粒。在此过程中,宿主细胞逐渐通过噬菌体酶削弱并最终破裂,平均释放100-200个新的噬菌体后代进入周围环境。

图2。描绘噬菌体裂解周期的阶段。
观看行动中的裂解周期这里。
溶菌发生循环
溶菌发生循环(图3)有时称为温带或非流动性感染,不会杀死宿主细胞,而是将其用作处于休眠状态的避难所。将噬菌体DNA注入宿主细胞后,它在噬菌体编码的积分酶的帮助下将自身整合到宿主基因组中,然后将其称为预言。然后,预言基因组与宿主基因组一起被动地复制,因为宿主细胞会划分,只要它留在那里,并且不会形成产生后代所需的蛋白质。由于噬菌体基因组通常相对较小,因此细菌宿主通常对此过程相对不受伤害。

图3。描绘噬菌体溶菌发生循环的阶段。
从溶菌发生到裂解的过渡
如果含有预言的细菌暴露于压力源,例如紫外线,养分低或丝裂霉素C等化学物质,则预言可能会自发地从宿主基因组中自发提取自己,并在称为诱导的过程中输入裂解周期。
但是,这个过程并不完美,预言有时可能会在重新循环时将其部分DNA留在后面,或者将部分宿主DNA随身携带。如果他们感染了新的宿主细胞,则可以在称为转导的过程中将细菌基因从一种菌株传输到另一种菌株。这是一种方法,抗生素抗性基因,毒素和超抗原编码基因以及其他毒力性状可能会通过细菌种群传播。
最近的工作表明,裂解和溶菌发生感染之间的过渡也取决于某个区域中的噬菌体丰度,因为它们能够在类似于法规传感的过程中产生并感知小肽4。
细菌免疫对噬菌体感染
并非所有细菌都对噬菌体攻击无助,拥有“免疫系统”,使他们能够反击。CRISPR-CAS现在是遗传修饰的代名词,首先提出了弗朗西斯科·莫吉卡(Francisco Mojica)作为细菌的“适应性免疫系统”5并由巴黎大学的一群人独立6在2005年。CRISPR基因座是一个短的重复序列的数组,该序列被带有独特序列的间隔者隔开。发现这些间隔序列与病毒和质粒DNA(包括噬菌体)具有同源性。当受到以前未经遇到的噬菌体的攻击时,在CRISPR的一侧添加了新的垫片,这使CRISPR成为了细胞及其祖先遇到的噬菌体的时间顺序记录。为了响应噬菌体的侵袭,CRISPR序列被转录,并与CAS蛋白合作,靶向并破坏与间隔者序列同源的噬菌体序列。
噬菌体作为遗传和分子生物学工具
lambda噬菌体,最初与大肠杆菌,是研究噬菌体最好的噬菌体之一,是许多遗传工具的基础。甚至有人说,将噬菌体用作工具最终导致了分子生物学的发展7。在1950年代,首先利用噬菌体与宿主DNA重组的能力来操纵基因组沙门氏菌物种,因此转导出生于8。从那时起,它一直用作在许多生物之间移动遗传物质的载体,包括真菌基因操纵9甚至人类基因。这要归功于谦虚的噬菌体人类胰岛素首先是安全廉价生产的。它还为高吞吐量筛选,纳米材料开发开放了应用程序10,,,,食品的抗菌治疗, 作为一个诊断工具以及药物比利时罗马尼亚比分直播发现和输送系统11。
噬菌体ϕx174在1977年成为不知不觉的先驱,当时它是第一个通过Fred Sanger和同事确定其整个核苷酸序列的生物12。
噬菌体疗法
在亚历山大·弗莱明(Alexander Fleming)在1928年发现抗生素之前,正在研究噬菌体作为治疗细菌感染的一种方法。在抗生素后时代,抗生素治疗的方便广谱活动意味着在大多数组织中对噬菌体疗法的研究被放弃。但是,在许多缺乏西方抗生素的前苏联国家中,对噬菌体疗法的研究继续通过必要。随着越来越多的全球问题抗生素耐药性,近年来,噬菌体疗法领域的复活。虽然噬菌体能够感染和破坏细菌,并已成功地用于治疗生命的感染13,它们的物种,甚至是应变特异性以及某些细菌预先存在免疫力的潜力,意味着针对噬菌体治疗的物种目前并不是一个微不足道的过程,必须针对个体感染量身定制。这使其昂贵且冗长。因此,目前这是最后的手段,在该领域仍然需要很多工作。
噬菌体家谱
随着核苷酸测序的可用性和负担能力的增加,在过去的二十年中,噬菌体基因组的数量爆炸了14。
噬菌体由国际病毒分类学委员会(ICTV)截至2017年更新,有19个噬菌体家庭感染细菌和古细菌(表1),但是随着来自较偏远地区的越来越多的样本被测序,这只会在将来生长。
对于移动用户,左右滚动以查看下面的表数据。
| 命令 | 家庭 | 形态学 | 核酸 | 例子 | 亚家族 | 属 |
| 尾病毒 | Ackermannviridae | dsdna | 2 | 4 | ||
| Myoviridae | 未发达的收缩尾巴 | 线性dsDNA | T4噬菌体,MU,PBSX,P1PUNA,P2,I3,BCEP 1,BCEP 43,BCEP 78 | 6 | 41 | |
| siphoviridae | 非共同的,非收集尾巴(长) | 线性dsDNA | λ噬菌体,T5噬菌体,Phi,C2,L5,HK97,N15 | 11 | 100 | |
| Podoviridae | 非共同的,非收集尾巴(短) | 线性dsDNA | T7噬菌体,T3噬菌体,φ29,p22,p37 | 3 | 23 | |
| Ligamenvirales | Lipothrixviridae | 包裹,杆状 | 线性dsDNA | 酸性丝状病毒1 | 3 | |
| Rudiviridae | 非发电,杆状 | 线性dsDNA | 磺胺岛岛棒状病毒1 | 1 | ||
| 未分配 | ampullaviridae | 包裹,瓶形 | 线性dsDNA | 1 | ||
| Bicaudaviridae | 未发育,柠檬形 | 圆形DSDNA | 1 | |||
| clavaviridae | 非发电,杆状 | 圆形DSDNA | 1 | |||
| Corticoviridae | 非发电,等距 | 圆形DSDNA | 1 | |||
| 囊状病毒科 | 包裹,球形 | 分段dsRNA | 1 | |||
| Fuselloviridae | 未发育,柠檬形 | 圆形DSDNA | 2 | |||
| Globuloviridae | 包裹,等距 | 线性dsDNA | 1 | |||
| Guttaviridae | 非发电,卵形 | 圆形DSDNA | 2 | |||
| Inoviridae | 非发电,丝状 | 圆形ssDNA | M13 | 7 | ||
| Leviviridae | 非发电,等距 | 线性ssRNA | MS2,Qβ | 2 | ||
| 微囊病 | 非发电,等距 | 圆形ssDNA | φx174 | 2 | 6 | |
| plasmaviridae | 包裹,多形 | 圆形DSDNA | 1 | |||
| Tectiviridae | 非发电,等距 | 线性dsDNA | 2 |
表格1。ICTV噬菌体感染细菌和古细菌的分类分类。
参考
1。Twort FW。对超微观病毒性质的研究。柳叶刀。1915; 186(4814):1241-1243。doi:10.1016/S0140-6736(01)20383-3
2。D'Herelle F.在不可见的微生物对源自牙杆菌的拮抗作用中:F。D’Herelle先生的简短笔记,由Roux先生提出。1917年。Res Microbiol。2007; 158(7):553-554。doi:10.1016/j.resmic.2007.07.005
3。Taylor NMI,Prokhorov NS,Guerrero-Ferreira RC等。T4底板的结构及其在触发护套收缩中的功能。自然。2016; 533(7603):346-352。doi:10.1038/nature17971
4。Erez Z,Steinberger-Levy I,Shamir M等。病毒之间的通信指导裂解 - 赖以生殖力的决定。自然。2017; 541(7638):488-493。doi:10.1038/nature21049
6。Pourcel C,Salvignol G,Vergnaud G.耶尔森氏菌的CRISPR元素通过优先吸收噬菌体DNA获得新的重复,并为进化研究提供了其他工具。 微生物学(阅读) 。2005; 151(PT 3):653-663。doi: 10.1099/MIC.0.27437-0
9。Chaveroche MK,Ghigo JM,D'EnfertC。一种在丝状真菌曲霉中有效替代基因的快速方法。 核酸res 。2000; 28(22):e97。doi: 10.1093/nar/28.22.e97
10。Chung WJ,Sena M,Merzlyak A,Lee SW。2.206-噬菌体作为功能性纳米材料开发的工具。在:Ducheyne P,编辑。 全面的生物材料 。Elsevier;2011:95-111。doi: 10.1016/B978-0-08-055294-1.00064-7
13。Schooley RT,Biswas B,Gill JJ等。开发和使用个性化的基于噬菌体的治疗鸡尾酒,以治疗患者患有耐药性的鲍曼尼杆菌感染的患者。 抗菌剂化学者 。2017; 61(10):E00954-17。doi: 10.1128/aac.00954-17


